No edit summary |
|||
| Line 144: | Line 144: | ||
[[Category:Ion channels]] |
[[Category:Ion channels]] |
||
[[Category:Ionotropic receptors]] |
[[Category:Ionotropic receptors]] |
||
| + | [[Category:Nicotinic receptors]] |
||
[[Category:Neurotransmitters]] |
[[Category:Neurotransmitters]] |
||
[[Category:Autoantigens]] |
[[Category:Autoantigens]] |
||
Latest revision as of 08:19, 23 January 2009
Assessment |
Biopsychology |
Comparative |
Cognitive |
Developmental |
Language |
Individual differences |
Personality |
Philosophy |
Social |
Methods |
Statistics |
Clinical |
Educational |
Industrial |
Professional items |
World psychology |
Biological: Behavioural genetics · Evolutionary psychology · Neuroanatomy · Neurochemistry · Neuroendocrinology · Neuroscience · Psychoneuroimmunology · Physiological Psychology · Psychopharmacology (Index, Outline)
 |
 |
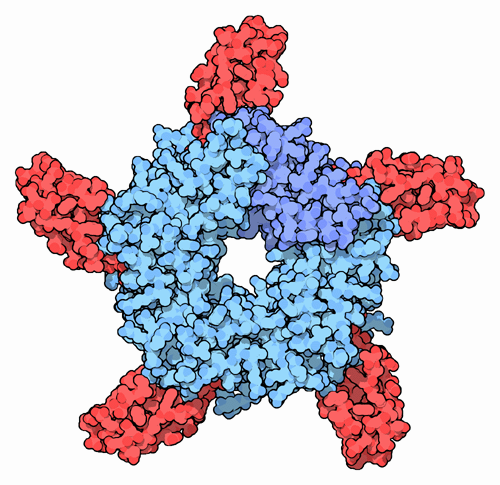
 Acetylcholine receptor (nicotinic) from electric torpedo rays (very similar to human receptor) is made of 5 subunits, 2 of which (shown in orange) bind ACh (red). Structure was determined by electron crystallography at 4 Å resolution (PDB code: 2bg9) (more details...) |
  Acetylcholine receptor blocked by cobra venom (PDB code: 1yi5). A similar effect can be achieved by high doses of curare or nicotine (more details...) |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Nicotinic acetylcholine receptors, or nAChRs, are cholinergic receptors that form ligand-gated ion channels in the plasma membranes of certain neurons. Being ionotropic (i.e. ligand-gated) receptors, nAChRs are directly linked to an ion channel and do not make use of a second messenger like metabotropic receptors do.[1]
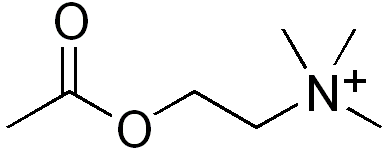
Like the other type of acetylcholine receptors - muscarinic acetylcholine receptors (mAChRs) - the opening of the ion pore of an nAChR is triggered by the binding of the neurotransmitter acetylcholine (ACh). However, whereas muscarinic receptors are also opened by muscarine, nicotinic receptors are also opened by nicotine. Hence, the name "nicotinic".[2][3][1]
Nicotinic acetylcholine receptors are present in many tissues in the body, and are the best-studied of the ionotropic receptors.[1] The neuronal receptors are found in the central nervous system and the peripheral nervous system. The neuromuscular receptors are found in the neuromuscular junctions of somatic muscles; stimulation of these receptors causes muscular contraction.
Structure
Nicotinic receptors, with a molecular mass of 290 kDa[4], are made up of five subunits, arranged symmetrically around the central pore.[1] They share similarities with GABAA receptors, glycine receptors, and the type 3 serotonin receptors (which are all ionotropic receptors), or the signature Cys-loop proteins.[5]
Twelve types of nicotinic receptor subunits, α2 through 10 and β2 through 4, combine to form pentamers. The subunits are somewhat similar to one another, especially in the hydrophobic regions. At the neuromuscular junction, the two α subunits of the nAChR are combined with up to four other subunits (β,γ,δ,ε) in the ratio 2α:β:ε:δ.[2][3][6][1] The neuronal forms are much more homogeneous, and are made up of only receptor subunit types (α and β) present in a ratio of 3α:2β. The neuronal forms also differ from the muscle forms in that they are not sensitive to α-bungarotoxin.[1]
The acetylcholine binding site is on the outside of the α subunit near the N terminus.[2] When an agonist binds to the site, the α subunits become more similar to the other subunits, the channel becomes more symmetrical,[7] and a pore with a diameter of about 0.65 nm opens.[2]
Opening the channel
Nicotinic AChRs may exist in different interconvertible conformational states. Binding of an agonist stabilizes the open and desensitised states. Opening of the channel allows positively charged ions to move across it, in particular, sodium and potassium, to enter the cell.
The nAChR is a non-selective cation channel, meaning that several different positively charged ions can cross through.[1] It is permeable to Na+ and K+, with some subunit combinations that are also permeable to Ca2+.[2] The amount of sodium and potassium the channels allow through their pores (their conductance) varies from 50-110 pS, with the conductance depending on the specific subunit composition as well as the permeant ion.[8]
Interestingly, because some neuronal nAChRs are permeable to Ca2+, they can affect the release of other neurotransmitters.[3] The channel usually opens rapidly and tends to remain open until the agonist diffuses away, which usually takes about 1 millisecond.[2] However, AChRs can sometimes open with only one agonist bound and in rare cases with no agonist bound, and they can close spontaneously even when ACh is bound. Therefore, ACh binding only creates a probability of pore opening, which increases as more ACh binds.[7]
Effects
This activation of receptors by nicotine modifies the state of neurons through two main mechanisms. On one hand, the movements of cations cause a depolarization of the plasma membrane (which results in an excitatory postsynaptic potential in neurons), but also by the activation of other voltage-gated ion channels. On the other hand, the entry of calcium acts, either directly or indirectly, on different intracellular cascades leading, for example, to the regulation of the activity of some genes or the release of neurotransmitters.
Receptor regulation
Receptor desensitization
Ligand-bound desensitization of receptors was first characterized by Katz and Thesleff in the nicotinic acetylcholine receptor[9]
Prolonged or repeat exposure to a stimulus often results in decreased responsiveness of that receptor for a stimulus. nAChR function can be modulated by phosphorylation[10] by the activation of second messenger-dependent protein kinases. Phosphorylation of the nAChR by PKA[9] and PKC[11] have been shown to phosphorylate nAChR resulting in its desensitization. It has been reported that after prolonged receptor exposure to the agonist, the agonist itself causes an agonist-induced conformational change in the receptor, resulting in receptor desensitization.[12] This receptor desensitization has been previously modeled in the context of a two-state mathematical model (see this link [1])
Roles
The subunits of the nicotinic receptors belong to a multigene family (17 members in human) and the assembly of combinations of subunits results in a large number of different receptors (For more information see the Ligand-Gated Ion Channel database). These receptors, with highly variable kinetic, electrophysiological and pharmacological properties, respond differently to nicotine, at very different effective concentrations. This functional diversity allows them to take part in two major types of neurotransmission. Classical synaptic transmission (wiring transmission) involves the release of high concentrations of neurotransmitter, acting on immediately neighbouring receptors. In contrast, paracrine transmission (volume transmission) involves neurotransmitters released by synaptic buttons, which then diffuse through the extra-cellular medium until they reach their receptors, which may be distant. Nicotinic receptors can also be found in different synaptic locations, for example the muscle nicotinic receptor always functions post-synaptically. The neuronal forms of the receptor can be found both post-synaptically (involved in classical neurotransmission) and pre-synaptically [13] where they can influence the release of multiple neurotransmitters.
Subunits
To date 17 nAChR subunits have been identified, these are divided into muscle-type and neuronal-type subunits. Of these 17 subunits, α2-α7 and β2-β4 have been cloned in humans, the remaining genes identified in chick and rat genomes.[14]
The nAChR subunits have been divided into 4 subfamilies (I-IV) based on similarities in protein sequence.[15] In addition, subfamily III has been further divided into 3 tribes.
| Neuronal-type | Muscle-type | ||||
| I | II | III | IV | ||
|---|---|---|---|---|---|
| α9, α10 | α7, α8 | 1 | 2 | 3 | α1, β1, δ, γ, ε |
| α2, α3, α4, α6 | β2, β4 | β3, α5 | |||
- Alpha genes: CHRNA1 (muscle), CHRNA2 (neuronal), CHRNA3, CHRNA4, CHRNA5, CHRNA6, CHRNA7, CHRNA8, CHRNA9, CHRNA10
- Beta genes: CHRNB1 (muscle), CHRNB2 (neuronal), CHRNB3, CHRNB4,
- Other genes: CHRND (delta), CHRNE (epsilon), CHRNG (gamma)
Notable variations
Nicotinic receptors are pentamers of these subunits, i.e. each receptor contains five subunits. Thus, there is an immense potential of variation of the aforementioned subunits. However, some of them are more notable than others, specifically (α1)2β1δε (muscle type), (α3)2(β4)3 (ganglion type), (α4)2(β2)3 (CNS type) and (α7)5 (another CNS type).[16] A comparison follows:
| Receptor type | Location | Effect | Nicotinic agonists | Antagonists |
|---|---|---|---|---|
| Muscle type: (α1)2β1δε[16] or α1β1δγε[17] |
Neuromuscular junction | EPSP, mainly by increased Na+ and K+ permeability |
| |
| Ganglion type: (α3)2(β4)3 |
autonomic ganglia | EPSP, mainly by increased Na+ and K+ permeability |
|
|
| CNS type: (α4)2(β2)3 |
Brain | Post- and presynaptic excitation,[16] mainly by increased Na+ and K+ permeability |
|
|
| (another) CNS type: (α7)5 |
Brain | Post- and presynaptic excitation,[16] mainly by increased Ca2+ permeability |
|
|
References
- ↑ 1.00 1.01 1.02 1.03 1.04 1.05 1.06 1.07 1.08 1.09 1.10 1.11 1.12 Purves, Dale, George J. Augustine, David Fitzpatrick, William C. Hall, Anthony-Samuel LaMantia, James O. McNamara, and Leonard E. White (2008). Neuroscience. 4th ed., 122-6, Sinauer Associates. ISBN 978-0-87893-697-7.
- ↑ 2.0 2.1 2.2 2.3 2.4 2.5 Siegel G.J., Agranoff B.W., Fisher S.K., Albers R.W., and Uhler M.D. (1999). Basic Neurochemistry: Molecular, Cellular and Medical Aspects (6th ed.). GABA Receptor Physiology and Pharmacology. American Society for Neurochemistry. URL accessed on 2008-10-01.
- ↑ 3.0 3.1 3.2 Itier V, Bertrand D (August 2001). Neuronal nicotinic receptors: from protein structure to function. FEBS letters 504 (3): 118–25.
- ↑ Unwin N. (March 2005). Refined structure of the nicotinic acetylcholine receptor at 4A resolution. Journal of Molecular Biology 346 (4): 967-89.
- ↑ Cascio, M. (May 2004). Structure and function of the glycine receptor and related nicotinicoid receptors. Journal of Biological Chemistry 279 (19): 19383-6.
- ↑ Giniatullin R, Nistri A, Yakel JL (July 2005). Desensitization of nicotinic ACh receptors: shaping cholinergic signaling. Trends Neurosci. 28 (7): 371-8.
- ↑ 7.0 7.1 Colquhoun D, Sivilotti LG. (June 2004). Function and structure in glycine receptors and some of their relatives. Trends Neurosci. 27 (6): 337-44.
- ↑ Mishina M, Takai T, Imoto K, Noda M, Takahashi T, Numa S, Methfessel C, Sakmann B (May 1986). Molecular distinction between fetal and adult forms of muscle acetylcholine receptor. Nature 321 (6068): 406-11.
- ↑ 9.0 9.1 Pitchford S, Day JW, Gordon A, Mochly-Rosen D (November 1992). Nicotinic acetylcholine receptor desensitization is regulated by activation-induced extracellular adenosine accumulation. The Journal of neuroscience : the official journal of the Society for Neuroscience 12 (11): 4540–4.
- ↑ Huganir RL, Greengard P (February 1983). cAMP-dependent protein kinase phosphorylates the nicotinic acetylcholine receptor. Proceedings of the National Academy of Sciences of the United States of America 80 (4): 1130–4.
- ↑ Safran A, Sagi-Eisenberg R, Neumann D, Fuchs S (August 1987). [hhttp://www.jbc.org/cgi/content/abstract/262/22/10506 Phosphorylation of the acetylcholine receptor by protein kinase C and identification of the phosphorylation site within the receptor delta subunit]. The Journal of biological chemistry 262 (22): 10506–10.
- ↑ Barrantes FJ (September 1978). Agonist-mediated changes of the acetylcholine receptor in its membrane environment. Journal of molecular biology 124 (1): 1–26.
- ↑ Wonnacott S (February 1997). Presynaptic nicotinic ACh receptors. Trends in neurosciences 20 (2): 92–8.
- ↑ Graham A, Court JA, Martin-Ruiz CM, Jaros E, Perry R, Volsen SG, Bose S, Evans N, Ince P, Kuryatov A, Lindstrom J, Gotti C, Perry EK (2002). Immunohistochemical localisation of nicotinic acetylcholine receptor subunits in human cerebellum. Neuroscience 113 (3): 493–507.
- ↑ Le Novère N, Changeux JP (February 1995). Molecular evolution of the nicotinic acetylcholine receptor: an example of multigene family in excitable cells. Journal of molecular evolution 40 (2): 155–72.
- ↑ 16.0 16.1 16.2 16.3 Rang, H. P. (2003). Pharmacology, 5th edition, Edinburgh: Churchill Livingstone.
- ↑ 17.0 17.1 17.2 Neurosci.pharm - MBC 3320 Acetylcholine
External links
| http://www.wikicities.com/images/Smallwikipedialogo.png | This page uses content from the English-language version of Wikiversity. The original article was at Poisson–Boltzmann profile for an ion channel. The list of authors can be seen in the page history. As with Psychology Wiki, the text of Wikiversity is available under the GNU Free Documentation License. |
Template:Autoantigens
Ion channel, receptor: ligand-gated ion channels | |
|---|---|
| Cys-loop receptors |
5-HT receptor (5-HT3 serotonin receptor (A)) - GABA receptor (GABA A (α1, α2, α3, α4, α5, α6, β1, β2, β3, γ2, γ3, ε), GABA C (ρ1, ρ2)) - Glycine receptor (α1, α2, β) - Nicotinic acetylcholine receptor (α1, α2, α3, α4, α5, α7, α9, β1, β2, β3, β4, δ, ε, (α4)2(β2)3, (α7)5, Ganglion type, Muscle type) |
| Ionotropic glutamate receptors |
AMPA (1, 2, 3, 4) - Kainate (1, 2, 3, 5) - NMDA (1, 2A, 2B, 2C, 2D) |
| ATP-gated channels |
Purinergic receptors (P2X (1, 2, 3, 4, 5, 7, L1)) |
| This page uses Creative Commons Licensed content from Wikipedia (view authors). |