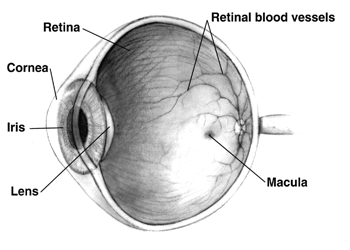
 Diagram of a human eye. Note that not all eyes have the same anatomy as a human eye. |
 Closeup of a blue-green human eye |
 The human eyes are sometimes metaphorically called "the windows to the soul." |
{kind=link}
{kind=link}
{kind=link}
An eye is an organ that detects light. Different kinds of light-sensitive organs are found in a variety of creatures. The simplest eyes do nothing but detect whether the surroundings are light or dark. More complex eyes are used to provide the sense of vision. Many complex organisms, including some mammal, birds, reptiles and fish, have two eyes which may be placed on the same plane to be interpreted as a single three-dimensional "image" (binocular vision), as in humans; or on different planes producing two separate "images" (monocular vision), such as in rabbits and chameleons.
Varieties of eyes

{kind=link}
Human eye


{kind=link}
The compound eyes of a dragonfly


{kind=link}
Compound eye of Antarctic krill
Assessment |
Biopsychology |
Comparative |
Cognitive |
Developmental |
Language |
Individual differences |
Personality |
Philosophy |
Social |
Methods |
Statistics |
Clinical |
Educational |
Industrial |
Professional items |
World psychology |
Biological: Behavioural genetics · Evolutionary psychology · Neuroanatomy · Neurochemistry · Neuroendocrinology · Neuroscience · Psychoneuroimmunology · Physiological Psychology · Psychopharmacology (Index, Outline)
In most vertebrates and some mollusks the eye works by allowing light to enter it and project onto a light-sensitive panel of cells known as the retina at the rear of the eye, where the light is detected and converted into electrical signals, which are then transmitted to the brain via the optic nerve. Such eyes are typically roughly spherical, filled with a transparent gel-like substance called the vitreous humour, with a focusing lens and often an iris which regulates the intensity of the light that enters the eye. The eyes of cephalopods, fish, amphibians, and snakes usually have fixed lens shapes, and focusing vision is achieved by telescoping the lens (similar to how a camera focuses).
Compound eyes are found among the arthropods and are composed of many simple facets which give a pixelated image (not multiple images as is often believed). Each sensor has its own lens and photosensitive cell(s). Some eyes have up to 28,000 such sensors, which are arranged hexagonally, and which can give a full 360 degree field of vision. Compound eyes are very sensitive to motion. Some arthropods (many Strepsiptera) have compound eye composed of a few facets each with a retina capable of creating an image, which does provide multiple image vision. With each eye viewing a different angle, a fused image from all the eyes is produced in the brain providing a very wide angle high resolution image.
Trilobites, which are now extinct, had unique compound eyes. They used clear calcite crystals to form the lenses of their eyes. In this, they differ from most other arthropods, which have soft eyes. The number of lenses in such an eye varied, however: some trilobites had only one, and some had thousands of lenses in one eye.
Some of the simplest eyes, called ocelli, can be found in animals like snails, who can not actually "see" in the common sense. They do have photosensitive cells, but no lens and no other means of projecting an image onto these cells. They can distinguish between light and dark (day and night), but no more. This enables snails to keep out of direct sunlight. Jumping spiders have simple eyes that are so large, supported by an array of other smaller eyes, that they can get enough visual inputs to hunt and pounce on their prey. Some insect larvae like caterpillars have a different type of single eye (stemmata) which gives a rough image.
Evolution of eyes
How a complex structure like the projecting eye could have evolved is often said to be a difficult question for the theory of evolution. Darwin famously treated the subject of eye evolution in his Origin of Species:
- To suppose that the eye, with all its inimitable contrivances for adjusting the focus to different distances, for admitting different amounts of light, and for the correction of spherical and chromatic aberration, could have been formed by natural selection, seems, I freely confess, absurd in the highest possible degree. Yet reason tells me, that if numerous gradations from a perfect and complex eye to one very imperfect and simple, each grade being useful to its possessor, can be shown to exist; if further, the eye does vary ever so slightly, and the variations be inherited, which is certainly the case; and if any variation or modification in the organ be ever useful to an animal under changing conditions of life, then the difficulty of believing that a perfect and complex eye could be formed by natural selection, though insuperable by our imagination, can hardly be considered real.
Despite the precision and complexity of the eye, theoretical analysis of eye evolution, developed by Dan-Erik Nilsson and Susanne Pelger (Nilsson and Pelger, 1994, Proc Biol Sci), demonstrated that a primitive optical sense organ could evolve into a complex human-like eye within a reasonable period (less than a million years) simply through small mutations and natural selection. Pro-intelligent design mathematician and Oxford professor David Berlinski [1] criticized these findings, including a criticism that the work contained no computer simulations (something assumed by a number of scientists but disclaimed by the original authors), and criticisms of the scientific establishment in general. [2] The original authors and other pro-evolution scientists subsequently challenged Berlinski's criticisms. [3]
Eyes in various animals show adaption to their requirements. For example, birds of prey have much greater visual acuity than humans and some, like diurnal birds of prey, can see ultraviolet light. The different forms of eye in, for example, vertebrates and mollusks are often cited as examples of parallel evolution. However, the development of the eye is considered by many experts to be monophyletic; that is, all modern eyes, varied as they are, have their origins in a proto-eye believed to have evolved some 540 million years ago (Mya).
As far as the vertebrate/mollusk eye is concerned, intermediate, functioning stages have existed in nature, which is also an illustration of the many varieties and peculiarities of eye construction. In the monophyletic model, these variations are less illustrative of non-vertebrate types such as the arthropod (compound) eye, but as those eyes are simpler to begin with, there are fewer intermediate stages to find.
- Eye Spot- A simple patch of photosensitive cells, common among lower invertebrates. Can sense ambient brightness. Physically similar to receptor patches for taste and smell. Some organisms cover the spot in optically-transparent skin cells.
- Pit Eye- The patch gradually depresses into a cup, which first grants the ability to discriminate brightness in directions, then in finer and finer directions as the pit deepens. Pit eyes were seen in ancient snails, and are found in some invertebrates living today.
- "Pinhole Camera" Eye- As the pit deepens into a cup, then a chamber, the opening of the chamber achieves true imaging, for fine directional sensing and some shape sensing. Currently found in the Nautilus.
- Pinhole Camera with Protective Layer- An overgrowth of transparent cells prevents contamination and parasitic infestation. The chamber contents, now segregated, can slowly specialize into a transparent humour, for optimizations such as color filtering, higher refractive index, blocking of ultraviolet, or the ability to operate in and out of water. The layer may in certain classes be related to the moulting of the organism's shell or skin.
- Multiple Humours- The transparent cells over the aperture split into two layers, with liquid in between. The liquid originally serves as a circulatory fluid for oxygen, nutrients, wastes, and immune functions, allowing greater total thickness and higher mechanical protection. In addition, multiple interfaces between solids and liquids increase optical power, allowing wider viewing angles, greater imaging resolution, or both. Again, division of layers may have originated with the shedding of skin; intracellular fluid may infill naturally depending on layer depth. Note: this layout has not been found, nor is it expected to be found. Fossilization rarely preserves soft tissues. In case it does, the new humour would almost certainly close as the remains dessicate, or as sediment overburden forces the layers together. Then the fossilized eye would resemble the previous layout.
- Crystalline Lens- It is biologically difficult to maintain a transparent layer of cells as sizes and thicknesses gradually increase. Deposition of transparent but nonliving material eases the need for nutrient supply and waste removal. In trilobites the material was calcite; in humans the material is a mixture of proteins called crystallins. A gap between tissue layers naturally forms a biconvex shape, which is optically and mechanically ideal for substances of normal refractive index. A biconvex lens confers not only optical resolution, but aperture and low-light ability, as resolution is now decoupled from hole size (which slowly increases again, free from the circulatory constraints).
- Separate Cornea and Iris- Independently, a transparent layer and a nontransparent layer may split forward from the lens. (These may happen before or after crystal deposition, or not at all.) Separation of the forward layer again forms a humour, the aqueous humour. This increases refractive power and again eases circulatory problems. Formation of a nontransparent ring allows more blood vessels, more circulation, and larger eye sizes. This flap around the perimeter of the lens also masks optical imperfections, which are more common at lens edges. The need to mask lens imperfections gradually increases with lens curvature and power, overall lens and eye size, and the resolution and aperture needs of the organism, driven by hunting or survival requirements. This type is now functionally identical to the eye of most vertebrates, including humans.
- "Backward" Illumination of Retina- The retina may revert on itself, forming a double layer. The nerves and blood vessels can migrate to the middle, where they do not block light, or form a blind spot on the retina. This type is seen in squids, which live in the dim oceans. In cats, which hunt at night, the retina does not revert. Instead a second, reflective layer (the tapetum) forms behind the retina. Light which is not absorbed by the retina on the first pass may bounce back and be detected. As a predator, the cat simply accommodates blind spots with head and eye motion.
At some point, color vision develops when receptor cells develop multiple pigments. As a chemical instead of mechanical adaptation, this may happen at any of the points described above, or not at all, and the capability may disappear and reappear, as organisms become predator or prey. Similarly, night and day vision emerge when receptors differentiate into rods and cones, respectively.
At some point, a focusing mechanism develops. Some species move the lens back and forth, some stretch the lens flatter. Another mechanism regulates focusing chemically and independently of these two, by controlling growth of the eye and maintaining focal length. Note that a focusing method is not a requirement. As photographers know, focal errors increase as f-number decreases. Thus, an organism with small eyes, active in direct sunlight, may survive with no focus mechanism at all. As the species grows larger, or transitions to dimmer environs, a means of focusing could appear gradually.
The majority of the process is believed to have taken only a few million years, as the first predator to gain true imaging would have touched off an "arms race." Prey animals and competing predators alike would be forced to rapidly match or exceed any such capabilities to survive. Hence multiple eye types and subtypes developed in parallel.
Anatomy


{kind=link}
Schematic diagram of the human eye.


{kind=link}
Light from a single point of a distant object and light from a single point of a near object being brought to a focus.
The structure of the mammalian eye owes itself completely to the task of focusing light onto the retina. All of the individual components through which light travels within the eye before reaching the retina are transparent, minimising dimming of the light. The cornea and lens help to converge light rays to focus onto the retina. This light causes chemical changes in the photosensitive cells of the retina, the products of which trigger nerve impulses which travel to the brain.
Light enters the eye from an external medium such as air or water, passes through the cornea, and into the first of two humours, the aqueous humour. Most of the light refraction occurs at the cornea which has a fixed curvature. The first humour is a clear mass which connects the cornea with the lens of the eye, helps maintain the convex shape of the cornea (necessary to the convergence of light at the lens) and provides the corneal endothelium with nutrients. The iris, between the lens and the first humour, is a coloured ring of muscle fibres. Light must first pass though the centre of the iris, the pupil. The size of the pupil is actively adjusted by the circular and radial muscles to maintain a relatively constant level of light entering the eye. Too much light being let in could damage the retina, too little light would be blinding. The lens, behind the iris, is a convex, springy disk which focuses light, through the second humour, onto the retina.
To clearly see an object far away, the circularly arranged ciliary muscles will pull on the lens, flattening it. Without muscles pulling on it, the lens will spring back into a thicker, more convex, form. Humans gradually lose this flexibility with age, resulting in the inability to focus on nearby objects, which is known as presbyopia. There are other refraction errors arising from the shape of the cornea and lens, and from the length of the eyeball. These include myopia, hyperopia, and astigmatism.
On the other side of the lens is the second humour, the vitreous humour, which is bounded on all sides: by the lens, ciliary body, suspensory ligaments and by the retina. It lets light through without refraction, helps maintain the shape of the eye and suspends the delicate lens.
Three layers, or tunics, form the wall of the eyeball. The outermost is the sclera which gives the eye most of its white colour. It consists of dense connective tissue filled with the protein collagen to both protect the inner components of the eye and maintain its shape. On the inner side of the sclera is the choroid, which contains blood vessels that supply the retinal cells with necessary oxygen and remove the waste products of respiration. Within the eye, only the sclera and ciliary muscles contain blood vessels. The choroid gives the inner eye a dark colour, which prevents disruptive reflections within the eye. The inner most layer of the eye is the retina, containing the photosensitive rod and cone cells, and neurons.
To maximise vision and light absorption, the retina is a relatively smooth (but curved) layer. It does have two points at which it is different; the fovea and blind spot. The fovea is a dip in the retina directly opposite the lens, which is densely packed with cone cells. It is largely responsible for colour vision in humans, and enables high acuity, such as is necessary in reading. The blind spot is a point on the retina where the optic nerve pierces the retina to connect to the nerve cells on its inside. No photosensitive cells exist at this point, it is thus "blind".
In some animals, the retina contains a reflective layer (the tapetum lucidum) which increases the amount of light each photosensitive cell perceives, allowing the animal to see better under low light conditions.
Other articles regarding eye anatomy
Aqueous humour, Anterior chamber, Blind spot, Ciliary body, Ciliary muscle, Cornea, Conjunctiva, Choroid, Fovea, Iris, Lens, Macula, Optic disc, Optic nerve, Ora serrata, Posterior chamber, Pupil, Retina, Schlemm's canal, Sclera, Suspensory ligament, Tapetum lucidum, Trabecular meshwork, Vitreous humour, Zonule of Zinn.
Cytology
The retina contains two forms of photosensitive cells - rods and cones. Though structurally and metabolically similar, their function is quite different, though they are equally important to vision. Rod cells are highly sensitive to light allowing them to respond in dim light and dark conditions. These are the cells which allow humans and other animals to see by moonlight, or with very little available light (as in a dark room). However, they do not distinguish between colours, and have low visual acuity (a measure of detail). This is why the darker conditions become, the less colour objects seem to have. Cone cells, conversely, need high light intensities to respond and have high visual acuity. Different cone cells respond to different colours (wavelengths) of light, which allows an organism to see colour.
The differences are useful; apart from enabling sight in both dim and light conditions, humans have given them further application. The fovea, directly behind the lens, consists of mostly densely-packed cone cells. This gives humans a highly detailed central vision, allowing reading, bird watching, or any other task which primarily requires looking at things. Its requirement for high intensity light does cause problems for astronomers, as they cannot see dim stars, or other objects, using central vision because the light from these is not enough to stimulate cone cells. Because cone cells are all that exist directly in the fovea, astronomers have to look at stars through the "corner of their eyes" (averted vision) where rods also exist, and where the light is sufficient to stimulate cells, allowing the individual to observe distant stars.
Rods and cones are both photosensitive, but respond differently to different frequencies of light. They both contain different pigmented photoreceptor proteins. Rod cells contain the protein rhodopsin and cone cells contain different proteins for each colour-range. The process through which these proteins go is quite similar - upon being subjected to electromagnetic radiation of a particular wavelength and intensity (ie. a colour visible light) the protein breaks down into two constituent products. Rhodopsin, of rods, breaks down into opsin and retinal; iodopsin of cones breaks down into photopsin and retinal. The opsin in both opens ion channels on the cell membrane which leads to the generation of an action potential (an impulse which will eventually get to the visual cortex in the brain).
This is the reason why cones and rods enable organisms to see in dark and light conditions - each of the photoreceptor proteins requires a different light intensity to break down into the constituent products. Further, synaptic convergence means that several rod cells are connected to a single bipolar cell, which then connects to a single ganglion cell and information is relayed to the visual cortex. Whereas, a single cone cell is connected to a single bipolar cell. Thus, action potentials from rods share neurons, where those from cones are given their own. This results in the high visual acuity, or the high ability to distinguish between detail, of cone cells and not rods. If a ray of light were to reach just one rod cell this may not be enough to stimulate an action potential. Because several "converge" onto a bipolar cell, enough transmitter molecules reach the synapse of the bipolar cell to attain the threshold level to generate an action potential.
Furthermore, colour is distinguishable when breaking down the iodopsin of cone cells because there are three forms of this protein. One form is broken down by the particular EM wavelength that is red light, another green light, and lastly blue light. In simple terms, this allows human beings to see red, green and blue light. If all three forms of cones are stimulated equally, then white is seen. If none are stimulated, black is seen. Most of the time however, the three forms are stimulated to different extents - resulting in different colours being seen. If, for example, the red and green cones are stimulated to the same extent, and no blue cones are stimulated, yellow is seen. For this reason red, green and blue are called primary colours and the colours obtained by mixing two of them, secondary colours. The secondary colours can be further complimented with primary colours to see tertiary colours.
Acuity
- Main article: Visual acuity
Visual acuity can be measured with several different metrics.
Cycles per degree (CPD) measures how much an eye can differentiate one object from another in terms of degree angles. It is essentially no different from angular resolution. To measure CPD, first draw a series of black and white lines of equal width on a grid (similar to a bar code). Next, place the observer at a distance such that the sides of the grid appear one degree apart. If the grid is 1 meter away, then the grid should be about 8.7 millimeters wide. Finally, increase the number of lines and decrease the width of each line until the grid appears as a solid grey block. In one degree, a human would not be able to distinguish more than about 12 lines without the lines blurring together. So a human can resolve distances of about 0.73 millimeters at a distance of one meter. A horse can resolve about 14 CPD (0.62 mm at 1 m) and a rat can resolve about 1 CPD (8.7 mm at 1 m).
A diopter is the unit of measure of focus.
Dynamic range
At any given instant, the retina can resolve a contrast ratio of around 100:1 (about 6 1/2 stops). As soon as your eye moves (saccades) it re-adjusts its exposure both chemically and by adjusting the iris. Initial dark adaptation takes place in approximately four seconds of profound, uninterrupted darkness; full adaptation through adjustments in retinal chemistry (the Purkinje effect) are mostly complete in thirty minutes. Hence, over time, a contrast ratio of about 1,000,000:1 (about 20 stops) can be resolved. The process is nonlinear and multifaceted, so an interruption by light nearly starts the adaptation process over again. Full adaptation is dependent on good blood flow; thus dark adaptation may be hampered by poor circulation, and vasoconstrictors like alcohol or tobacco.
The orbit
In many species, the eyes are inset in the portion of the skull known as the orbits or eyesockets. This placement of the eyes helps to protect them from injury.
Eyebrows
In humans, the eyebrows redirect flowing substances (usually rainwater) away from the eye. Water in the eye can alter the refractive properties of the eye and blur vision. It can also wash away the tear fluid - along with it the protective lipid layer - and can alter corneal physiology, due to osmotic differences between tear fluid and freshwater. This is made apparent when swimming in freshwater pools, as the osmotic gradient draws 'pool water' into the corneal tissue, causing edema, and subsequently leaving the swimmer with "cloudy" or "misty" vision for a short period thereafter. It can be reversed by irrigating the eye with hypertonic saline.
Eyelids
In many animals, including humans, eyelids wipe the eye and prevent dehydration. They spread tear fluid on the eyes, which contains substances which help fight bacterial infection as part of the immune system. Some aquatic animals have a second eyelid in each eye which refracts the light and helps them see clearly both above water and below it. Most creatures will automatically react to a threat to its eyes (such as an object moving straight at the eye, or a bright light) by covering the eyes, and/or by turning the eyes away from the threat. Blinking the eyes is, of course, also a reflex.
Eyelashes
In many animals, including humans, eyelashes prevent fine particles from entering the eye. Fine particles can be bacteria, but also simple dust which can cause irritation of the eye, and lead to tears and subsequent blurred vision.
Eye movement
- Main article: Eye movements
Animals with compound eyes have a wide field of vision, allowing them to look in many directions. To see more, they have to move their entire head or even body.
The visual system in the brain is too slow to process that information if the images are slipping across the retina at more than a few degrees per second (Westheimer and McKee, 1954). Thus, for humans to be able to see while moving, the brain must compensate for the motion of the head by turning the eyes. Another complication for vision in frontal-eyed animals is the development of a small area of the retina with a very high visual acuity. This area is called the fovea, and covers about 2 degrees of visual angle in people. To get a clear view of the world, the brain must turn the eyes so that the image of the object of regard falls on the fovea. Eye movements are thus very important for visual perception, and any failure to make them correctly can lead to serious visual disabilities. To see a quick demonstration of this fact, try the following experiment: hold your hand up, about one foot (30 cm) in front of your nose. Keep your head still, and shake your hand from side to side, slowly at first, and then faster and faster. At first you will be able to see your fingers quite clearly. But as the frequency of shaking passes about one hertz, the fingers will become a blur. Now, keep your hand still, and shake your head (up and down or left and right). No matter how fast you shake your head, the image of your fingers remains clear. This demonstrates that the brain can move the eyes opposite to head motion much better than it can follow, or pursue, a hand movement. When your pursuit system fails to keep up with the moving hand, images slip on the retina and you see a blurred hand. Having two eyes is an added complication, because the brain must point both of them accurately enough that the object of regard falls on corresponding points of the two retinas; otherwise, double vison would occur. The movements of different body parts are controlled by striated muscles acting around joints. The movements of the eye are no exception, but they have special advantages not shared by skeletal muscles and joints, and so are considerably different.
Where the parts of an eye are located
- Lens - Immediately behind the iris
- Cornea - The dome on the front of the eye
- Retina - Rear of the eye
- Pupil - Middle front of the eye
- Iris - Right around the pupil
- Optic nerve - Connects to the center of the retina
- Sclera - The outer part of the eye that forms the white around the pupil and iris
How we see an object
The steps of how we see an object: 1) Enters the cornea 2) Through the pupil 3) Through the Iris 4) Then Through the crystalline lens 5) Then Through the vitreous humor 6) Then Through the retina 7) Then Through the optic nerve 8) Then Through the visual pathway 9) Then Through the occipital cortex 10) Then Through the brain
How we see color
How we see color: 1) All the “invisible” colors of sun light shine on object 2) The surface of object absorb all colored light rays , except for those corresponding to the color of object, and reflects this color to the human eye 3) The eye receives the reflected color and sends a message to the brain
Extraocular muscles
- Main article: Extraocular muscles
Each eye has six muscles that control its movements: the lateral rectus, the medial rectus, the inferior rectus, the superior rectus, the inferior oblique, and the superior oblique. When the muscles exert different tensions, a torque is exerted on the globe that causes it to turn. This is an almost pure rotation, with only about one millimeter of translation (Carpenter, 1988). Thus, the eye can be considered as undergoing rotations about a single point in the center of the eye.
Rapid eye movement
- Main article: Rapid eye movement
Rapid eye movement typically refers to the stage during sleep during which the most vivid dreams occur. During this stage, the eyes move rapidly. It is not in itself a unique form of eye movement.
Saccades
- Main article: Saccade
Saccades are rapid refocussing actions of the eyes. Many animals are able to quickly look at a point in space (prompted by memory, peripheral vision or an audio cue) without actively looking at anything in between. The eyes simply jerk into a new position. Saccades move the eye at up to 900°/s in adult humans, and take roughly 250 milliseconds to be initiated by the neural network.
Microsaccades
- Main article: Microsaccade
Even when looking intently at a single spot, the eyes drift around. This ensures that individual photosensitive cells are continually stimulated in different degrees. Without changing input, these cells would otherwise stop generating output. Microsaccades move the eye no more than a total of 0.2° in adult humans.
Vestibulo-ocular reflex
- Main article: Vestibulo-ocular reflex
Many animals can look at something while turning their heads. The eyes are automatically rotated to remain fixed on the object, directed by input from the organs of balance near the ears.
Smooth pursuit movement
The eyes can also follow a moving object around. This is less accurate than the vestibulo-ocular reflex as it requires the brain to process incoming visual information and supply feedback. Following an object moving at constant speed is relatively easy, though the eyes will often make saccadic jerks to keep up. The smooth pursuit movement can move the eye at up to 100°/s in adult humans.
Optokinetic reflex
The optokinetic reflex is a combination of a saccade and smooth pursuit movement. When, for example, looking out of the window in a moving train, the eyes can focus on a 'moving' tree for a short moment (through smooth pursuit), until the tree moves out of the field of vision. At this point, the optokinetic reflex kicks in, and moves the eye back to the point where it first saw the tree (through a saccade).
Vergence movement
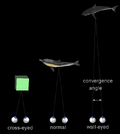
{kind=link}
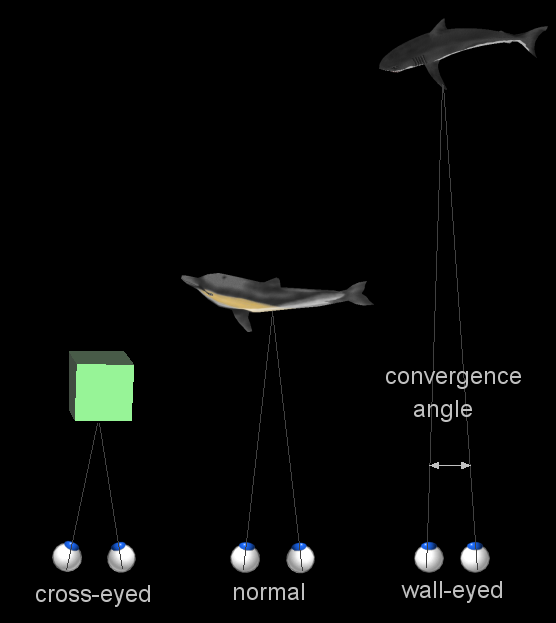
The two eyes converge to point to the same object
When a creature with binocular vision looks at an object, the eyes must rotate around a vertical axis so that the projection of the image is in the centre of the retina in both eyes. To look at an object closer by, the eyes rotate 'towards each other' (convergence), while for an object farther away they rotate 'away from eachother' (divergence). Exaggerated convergence is called cross eyed viewing (focussing on the nose for example) . When looking into the distance, or when 'staring into nothingness', the eyes neither converge nor diverge.
Vergence movements are closely connected to accommodation of the eye. Under normal conditions, changing the focus of the eyes to look at an object at a different distance will automatically cause vergence and accommodation.
Accommodation
- Main article: Accommodation (eye)
To see clearly, the lens will be pulled flatter or allowed to regain its thicker form.
Main articles: List of eye diseases and disorders and Ocular manifestations of systemic disease
There are many diseases and disorders that may affect the eyes.
As the eye ages certain changes occur that can be attributed to solely the aging process. Most of these anatomic and physiologic processes follow a gradual decline. With aging, the quality of vision worsens due to reasons independent of aging eye diseases. While there are many changes of significance in the nondiseased eye, the most functionally important changes seem to be a reduction in pupil size and the loss of accommodation or focusing capability (presbyopia). The area of the pupil governs the amount of light that can reach the retina. The extent to which the pupil dilates also decreases with age. Because of the smaller pupil size, older eyes receive much less light at the retina. In comparison to younger people, it is as though older persons wear medium-density sunglasses in bright light and extremely dark glasses in dim light. Therefore, for any detailed visually guided tasks on which performance varies with illumination, older person requires extra lighting. [4]
With aging a prominent white ring develops in the periphery of the cornea- called arcus senilis. Aging causes laxity and downward shift of eyelid tissues and atrophy of the orbital fat. These changes contribute to the etiology of several eyelid disorders such as ectropion, entropion, dermatochalasis, and ptosis. The vitreous gel undergoes liquefaction (posterior vitreous detachment or PVD) and its opacities - visible as floaters gradually increase in number.
See also
- WikiSaurus:eye — the WikiSaurus list of synonyms and slang words for eyes in many languages
- Adaptation
- Binocular vision
- Corrective lens
- Crystallin
- Evil eye
- Eye color
- Eye contact
- Eye tracking
- Eyeglass prescription
- Macropsia
- Micropsia
- Nictitating membrane
- Ocular tremor
- Ophthalmology
- Optician
- Optometry
- Persistence of vision
- Phosphenes
- Snellen chart
- Staring contest
- Tears
- Visual perception
External links
- DJO | Digital Journal of Ophthalmology
- Glossary of Eye Conditions
- Evolution of the Eye
- eye Topics
- Diagram of the eye
- Webvision. The organisation of the retina and visual system.
- VisionSimulations.com | Images and vision simulators of various diseases and conditions of the eye
References
- Anatomy. History of Ophthalmology. URL accessed on 23 April, 2005.
- Kandel ER, Schwartz JH, Jessell TM. Principles of Neural Science, 4th ed. McGraw-Hill, New York (2000). ISBN 0838577016
- Internet lecture on eye types in animal kindom
| Sensory system - Visual system - Eye - edit |
|---|
| Anterior chamber | Aqueous humour | Blind spot | Choroid | Ciliary body | Conjunctiva | Cornea | Iris | Lens | Macula | Optic disc | Optic fovea | Posterior chamber | Pupil | Retina | Schlemm's canal | Sclera | Tapetum lucidum | Trabecular meshwork | Vitreous humour |
| Sensory system - Visual system - edit |
|---|
| Eye | Optic nerve | Optic chiasm | Optic tract | Lateral geniculate nucleus | Optic radiation | Visual cortex |
ar:عين bg:Око ca:Ull cs:Oko cy:Llygad da:Øje de:Auge es:Ojo eo:Okulo fi:Silmä fr:Œil he:עין id:Mata io:Okulo is:Auga lb:A lt:Akis ms:Mata nl:Oog nn:auga pt:Olho ro:Ochi ru:Глаз sk:Oko sl:Oko sr:Око su:Panon sv:Öga th:ตา zh:眼睛 zh-min-nan:Ba̍k-chiu
| This page uses Creative Commons Licensed content from Wikipedia (view authors). |