Assessment |
Biopsychology |
Comparative |
Cognitive |
Developmental |
Language |
Individual differences |
Personality |
Philosophy |
Social |
Methods |
Statistics |
Clinical |
Educational |
Industrial |
Professional items |
World psychology |
Biological: Behavioural genetics · Evolutionary psychology · Neuroanatomy · Neurochemistry · Neuroendocrinology · Neuroscience · Psychoneuroimmunology · Physiological Psychology · Psychopharmacology (Index, Outline)
{kind=link}
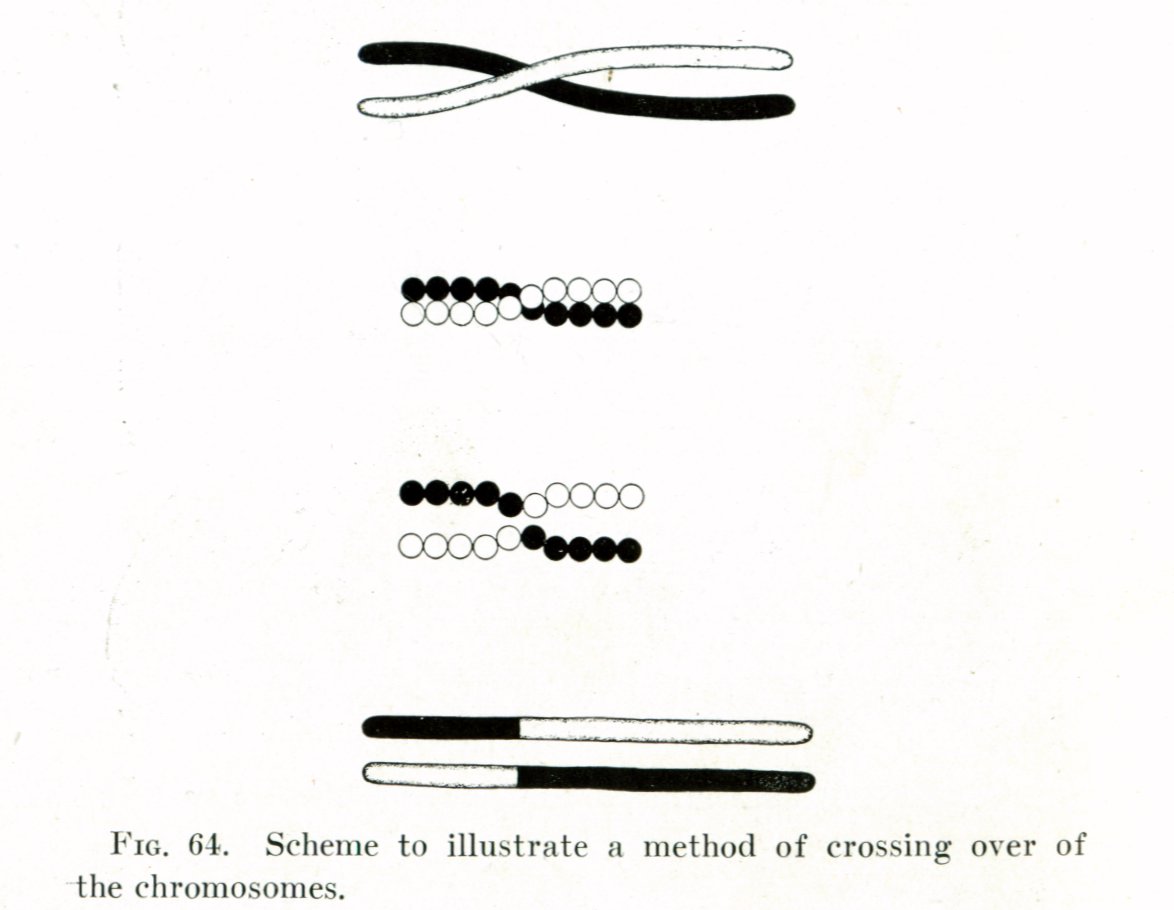
Crossing over occurs during meiosis I, and is the process where homologous chromosomes pair up with each other and exchange different segments of their genetic material to form recombinant chromosomes. Crossing over is essential for the normal segregation of chromosomes during meiosis. Crossing over also accounts for genetic variation, because due to the swapping of genetic material during crossing over, the chromatids held together by the centromere are no longer identical. So, when the chromosomes go on to meiosis II and separate, some of the daughter cells receive daughter chromosomes with recombined alleles. Due to this genetic recombination, the offspring have a different set of alleles and genes than their parents do. In the diagram, genes B and are a crossed over with each other, making the resulting recombinants after meiosis Ab, AB, ab, and aB.
{kind=link}
Thomas Hunt Morgan's illustration of crossing over (1916)
{kind=link}
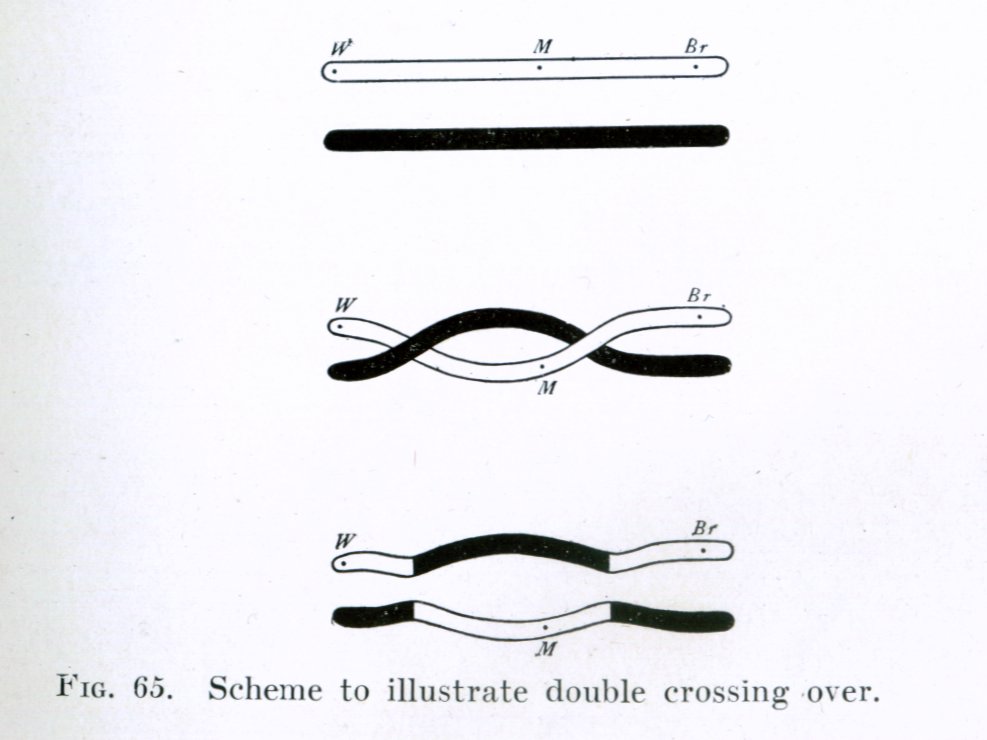
A double crossing over

{kind=link}
Recombination involves the breakage and rejoining of parental chromosomes
Chromosomal crossover (or crossing over) is the exchange of genetic material between homologous chromosomes that results in recombinant chromosomes. It is one of the final phases of genetic recombination, which occurs during prophase I of meiosis (pachytene) in a process called synapsis. Synapsis begins before the synaptonemal complex develops, and is not completed until near the end of prophase I. Crossover usually occurs when matching regions on matching chromosomes break and then reconnect to the other chromosome.
Crossing over was described, in theory, by Thomas Hunt Morgan. He relied on the discovery of the Belgian Professor Frans Alfons Janssens of the University of Leuven who described the phenomenon in 1909 and had called it 'chiasmatypie'. The term chiasma is linked if not identical to chromosomal crossover. Morgan immediately saw the great importance of Janssens' cytological interpretation of chiasmata to the experimental results of his research on the heredity of Drosophila. The physical basis of crossing over was first demonstrated by Harriet Creighton and Barbara McClintock in 1931.[1]
Chemistry[]
Meiotic recombination initiates with double-stranded breaks that are introduced into the DNA by the Spo11 protein.[2] One or more exonucleases then digest the 5’ ends generated by the double-stranded breaks to produce 3’ single-stranded DNA tails. The meiosis-specific recombinase Dmc1 and the general recombinase Rad51 coat the single-stranded DNA to form nucleoprotein filaments.[3] The recombinases catalyze invasion of the opposite chromatid by the single-stranded DNA from one end of the break. Next, the 3’ end of the invading DNA primes DNA synthesis, causing displacement of the complementary strand, which subsequently anneals to the single-stranded DNA generated from the other end of the initial double-stranded break. The structure that results is a cross-strand exchange, also known as a Holliday junction. The contact between two chromatids that will soon undergo crossing-over is known as a chiasma. The Holliday junction is a tetrahedral structure which can be 'pulled' by other recombinases, moving it along the four-stranded structure.
|
{kind=link}
{kind=link}
Consequences[]
{kind=link}
The difference between gene conversion and chromosomal crossover. Blue is the two chromatids of one chromosome and red is the two chromatids of another one.
In most eukaryotes, a cell carries two versions of each gene, each referred to as an allele. Each parent passes on one allele to each offspring. An individual gamete inherits a complete haploid complement of alleles on chromosomes that are independently selected from each pair of chromatids lined up on the metaphase plate. Without recombination, all alleles for those genes linked together on the same chromosome would be inherited together. Meiotic recombination allows a more independent selection between the two alleles that occupy the positions of single genes, as recombination shuffles the allele content between homologous chromosomes.
Recombination results in a new arrangement of maternal and paternal alleles on the same chromosome. Although the same genes appear in the same order, the alleles are different. In this way, it is theoretically possible to have any combination of parental alleles in an offspring, and the fact that two alleles appear together in one offspring does not have any influence on the statistical probability that another offspring will have the same combination. This theory of "independent assortment" of alleles is fundamental to genetic inheritance.[4] However, the frequency of recombination is actually not the same for all gene combinations. This leads to the notion of "genetic distance", which is a measure of recombination frequency averaged over a (suitably large) sample of pedigrees. Loosely speaking, one may say that this is because recombination is greatly influenced by the proximity of one gene to another. If two genes are located close together on a chromosome, the likelihood that a recombination event will separate these two genes is less than if they were farther apart. Genetic linkage describes the tendency of genes to be inherited together as a result of their location on the same chromosome. Linkage disequilibrium describes a situation in which some combinations of genes or genetic markers occur more or less frequently in a population than would be expected from their distances apart. This concept is applied when searching for a gene that may cause a particular disease. This is done by comparing the occurrence of a specific DNA sequence with the appearance of a disease. When a high correlation between the two is found, it is likely that the appropriate gene sequence is really closer.[5]
Problems[]
Although crossovers typically occur between homologous regions of matching chromosomes, similarities in sequence can result in mismatched alignments. These processes are called unbalanced recombination. Unbalanced recombination is fairly rare compared to normal recombination, but severe problems can arise if a gamete containing unbalanced recombinants becomes part of a zygote. The result can be a local duplication of genes on one chromosome and a deletion of these on the other, a translocation of part of one chromosome onto a different one, or an inversion.
References[]
- ↑ Creighton H, McClintock B (1931). A Correlation of Cytological and Genetical Crossing-Over in Zea Mays. Proc Natl Acad Sci USA 17 (8): 492–7. (Original paper)
- ↑ Keeney, S (1997). Meiosis-Specific DNA Double-Strand Breaks Are Catalyzed by Spo11, a Member of a Widely Conserved Protein Family. Cell 88 (3): 375–84.
- ↑ Sauvageau, S; Stasiak, Az; Banville, I; Ploquin, M; Stasiak, A; Masson, Jy (Jun 2005). Fission Yeast Rad51 and Dmc1, Two Efficient DNA Recombinases Forming Helical Nucleoprotein Filaments. Molecular and Cellular Biology 25 (11): 4377–87.
- ↑ genetic recombination.
- ↑ Genetic Recombination
See also[]
- Unequal crossing over
- Coefficient of coincidence
- Genetic distance
- Independent assortment
- Mitotic crossover
- Recombinant frequency
Template:Genetic recombination
| This page uses Creative Commons Licensed content from Wikipedia (view authors). |