Assessment |
Biopsychology |
Comparative |
Cognitive |
Developmental |
Language |
Individual differences |
Personality |
Philosophy |
Social |
Methods |
Statistics |
Clinical |
Educational |
Industrial |
Professional items |
World psychology |
Biological: Behavioural genetics · Evolutionary psychology · Neuroanatomy · Neurochemistry · Neuroendocrinology · Neuroscience · Psychoneuroimmunology · Physiological Psychology · Psychopharmacology (Index, Outline)
| Brain: Cingulate cortex | ||
|---|---|---|
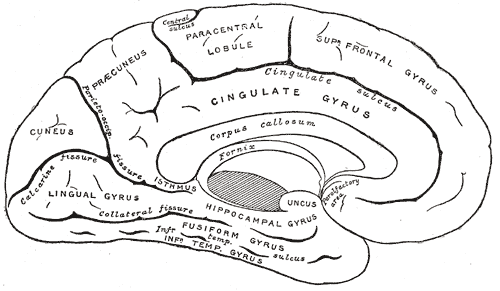 | ||
| Medial surface of left cerebral hemisphere. | ||
 | ||
| Medial surface. | ||
| Latin | gyrus cingulatus | |
| Gray's | subject #189 825 | |
| Part of | ||
| Components | ||
| Artery | ||
| Vein | ||
| BrainInfo/UW | hier-141 | |
| MeSH | A08.186.211.577.330 | |
{kind=link}
Sagittal MRI slice with highlighting indicating location of the cingulate cortex.
{kind=link}
Coronal section of brain. Cingulate cortex is shown in yellow.
The cingulate cortex is a part of the brain situated in the medial aspect of the cerebral cortex. It includes the cortex of the cingulate gyrus, which lies immediately above the corpus callosum, and the continuation of this in the cingulate sulcus. The cingulate cortex is usually considered part of the limbic lobe.
It receives inputs from the thalamus and the neocortex, and projects to the entorhinal cortex via the cingulum. It is an integral part of the limbic system, which is involved with emotion formation and processing, learning, and memory. The combination of these three functions makes the cingulate gyrus highly influential in linking behavioral outcomes to motivation (e.g. a certain action induced a positive emotional response, which results in learning).[1] This role makes the cingulate cortex highly important in disorders such as depression[citation needed] and schizophrenia. It also plays a role in executive function and respiratory control.
History[]
Cingulum means "belt" in Latin. The name was likely chosen because this cortex, in great part, surrounds the corpus callosum. Cingulate is an adjective (cingularis or cingulatus). The cingulate cortex is a part of the "grand lobe limbique" of Broca (1898) that consisted (in addition to the olfactory part, which is no more considered there today) of a superior cingulate part, supracallosal; and an inferior hippocampic part, infracallosal. The limbic lobe was separated from the remainder of the cortex by Broca for two reasons: first that it is not convoluted, and second that the gyri are directed parasagittally (contrary to the transverse gyrification). Since the parasagittal gyrification is observed in non-primate species, the limbic lobe was thus declared to be "bestial". As with other parts of the cortex, there have been and continue to be discrepancies concerning boundaries and naming. Brodmann (1909), a student of Cécile Vogt-Mugnier and Oskar Vogt, who worked on cercopithecus (and not much in human [Bailey and von Bonin]), elaborated a system of numeration that had unfortunately no typological logics (1, 2 and 3 are sensory, 4 is motor, 5 is parietal, 6 is premotor and 7 is again parietal!). Area 25 was even not placed by him in the same place in the human brain. Area 24 (anterior) was distinguished from 23 (posterior) on the basis that it was agranular. More recently, the typographical von Economo's system was adopted by Bailey and von Bonin. Simple typographical naming should be preferred, for evident heuristic purposes.
{kind=link}
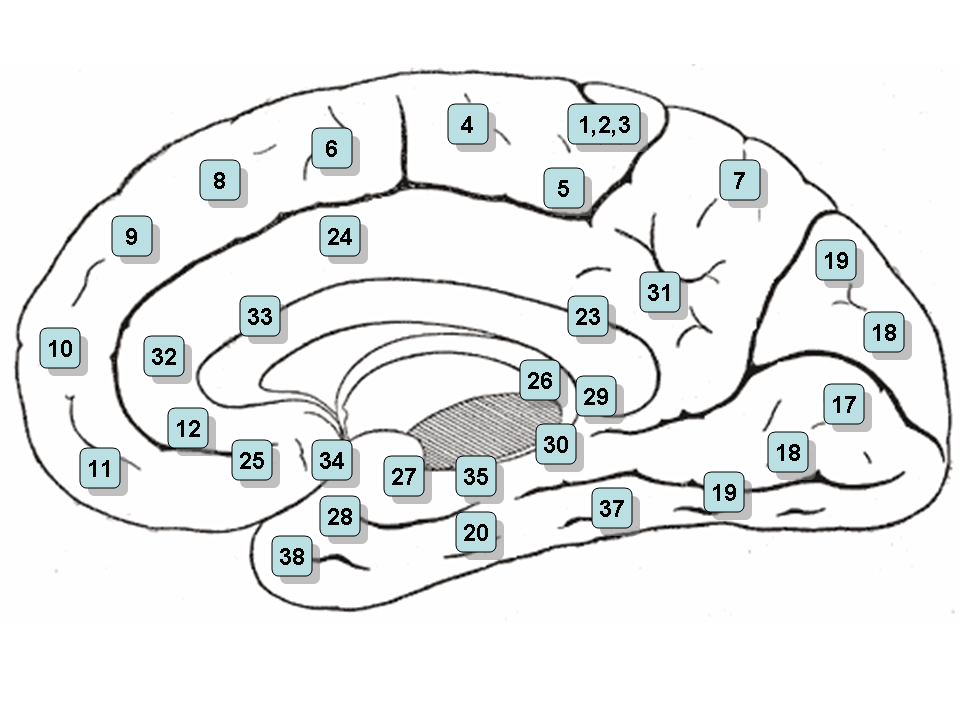
Brodmann areas of a medial section of the right hemisphere.
Subdivisions[]
Based on cerebral cytoarchitectonics it has been divided into the Brodmann areas 23, 24, 26, 29, 30, 31 and 32. The areas 26, 29 and 30 are usually referred to as the retrosplenial areas.
{kind=link}
Micrograph showing spindle neurons of the cingulate cortex. HE-LFB stain.
Anterior cingulate cortex[]
- Main article: Anterior cingulate cortex
This corresponds to areas 24, 32 and 33 of Brodmann and LA of Constantin von Economo and Bailey and von Bonin. It is continued anteriorly by the subgenual area (area 25, located below the genu of the corpus callosum). It is cytoarchitectonically agranular. It has a gyral and a sulcal part. Anterior cingulate cortex can further be divided in the perigenual anterior cingulate cortex (near the genu) and midcingulate cortex. The anterior cingulate cortex receives primarily its afferent axons from the intralaminar and midline thalamic nuclei (see thalamus). The nucleus anterior receives mamillo-thalamic afferences. The mamillary neurons receive axons from the subiculum. The whole forms a part of Papez' circuit. The anterior cingulate cortex sends axons to the anterior nucleus and through the cingulum to other Broca's limbic areas. The ACC is involved in error and conflict detection processes.
Posterior cingulate cortex[]
- Main article: Posterior cingulate cortex
This corresponds to area 23 of Brodmann LP of von Economo and Bailey and von Bonin. Its cellular structure is granular. It is followed posteriorly by the retrosplenial cortex (area 29).[citation needed] Dorsally is the granular area 31. The posterior cingulate cortex receives a great part of its afferent axons from the superficial nucleus (or nucleus superior- falsely LD-[citation needed]) of the thalamus (see thalamus), which itself receives axons from the subiculum. To some extent it thus duplicates Papez' circuit. It receives also direct afferents from the subiculum of the hippocampus. Posterior cingulate cortex hypometabolism (with 18F-FDG PET) has been defined in Alzheimer's disease.
Inputs of the anterior cingulate gyrus[]
A retrograde tracing experiment on macaque monkeys revealed that ventral anterior (VA) and ventral lateral (VL) nuclei of the thalamus are connected with motor areas of cingulate sulcus.[2] Retrosplenial region (Brodmann’s area 26, 29 and 30) part of cingulate gyrus can be divided into three parts retrosplenial granular cortex A, the retrosplenial granular cortex B and the retrosplenial dysgranular cortex. The hippocampal formation sends dense projections to the retrosplenial granular cortex A and B and fewer projections to the retrosplenial dysgranular cortex. Postsubiculum sends projections to the retrosplenial granular cortex A and B and to the retrosplenial dysgranular cortex. Dorsal subiculum sends projections to the retrosplenial granular cortex B, while ventral subiculum sends projections to the retrosplenial granular cortex A. Entorhinal cortex – caudal parts – sends projections to the retrosplenial dysgranular cortex.[3]
Outputs of the anterior cingulate gyrus[]
Rostral cingulate gyrus (Brodmanns’s area 32) projects to rostral superior temporal gyrus, midorbitofrontal cortex and lateral prefrontal cortex.[4] Ventral anterior cingulate (Brodmann’s area 24) sends projections to the anterior insular cortex, premotor cortex (Brodmann’s area 6), Brodmann’s area 8, the perirhinal area, the orbitofrontal cortex (Brodmann’s area 12), the laterobasal nucleus of amygdala, and the rostral part of the inferior parietal lobule.[4] Injecting wheat germ agglutinin and horseradish peroxidase conjugate into anterior cingulate gyrus of cats, revealed that anterior cingulate gyrus has reciprocal connections with rostral part of the thalamic posterior lateral nucleus and rostral end of the pulvinar.[5] Postsubiculum receives projections from the retrospleinal dysgranular cortex and the retrosplenial granular cortex A and B. Parasubiculum receives projections from the retrosplenial dysgranular cortex and retrosplenial granular cortex A. Caudal and lateral parts of the entorhinal cortex get projections from the retrosplenial dysgranular cortex, while caudal medial entorhinal cortex receives projections from the retrosplenial granular cortex A. The retrosplenial dysgranular cortex sends projections to the perirhinal cortex. The retrospleinal granular cortex A sends projection to the rostral presubiculum.[3]
Outputs of the posterior cingulate gyrus[]
The posterior cingulate cortex (Brodmann’s area 23) sends projections to dorsolateral prefrontal cortex (Brodmann’s area 9), anterior prefrontal cortex (Brodmann’s area 10), orbitofrontal cortex (Brodmanns’ area 11), the parahippocampal gyrus, posterior part of the inferior parietal lobule, the presubiculum, the superior temporal sulcus and the retrosplenial region.[4] The retrosplenial cortex and caudal part of the cingulate cortex are connected with rostral prefrontal cortex via cingulate fascicule in macaque monkeys[6] Ventral posterior cingulate cortex was found to be reciprocally connected with the caudal part of the posterior parietal lobe in rhesus monkeys.[7] Also the medial posterior parietal cortex is connected with posterior ventral bank of the cingulate sulcus.[7]
Other connections[]
The anterior cingulate is connected to the posterior cingulate at least in rabbits. Posterior cingulate gyrus is connected with retrosplenial cortex and this connection is part of the dorsal splenium of the corpus callosum. The anterior and posterior cingulate gyrus and retrosplenial cortex send projections to subiculum and presubiculum.[8]
Involvement in mental disorders[]
- See also: Anterior cingulate cortex
Schizophrenia[]
Using a three-dimensional magnetic resonance imaging procedure to measure the volume of the rostral anterior cingulate gyrus (perigenual cingulate gyrus), Takahashi et al. (2003) found that the rostral anterior cingulate gyrus is larger in control (healthy) females than males, but this sex difference was not found in schizophrenic patients. Schizophrenic patients also had a smaller volume of perigenual cingulate gyrus than control subjects.[9]
Haznedar et al. (2004) studied metabolic rate of glucose in anterior and posterior cingulate gyrus in patients with schizophrenia, schizotypal personality disorder (SPD) and compared them with a control group. The metabolic rate of glucose was found to be lower in the left anterior cingulate gyrus and the right posterior cingulate gyrus in patients with schizophrenia relative to controls. Although SPD patients were expected to show a glucose metabolic rate somewhere between the schizophrenic and controls, they actually had higher metabolic glucose rate in the left posterior cingulate gyrus. The volume of the left anterior cingulate gyrus was reduced in schizophrenic patients as compared with controls, but there was not any difference between SPD patients and schizophrenic patients. From these results it appears that the schizophrenia and SPD are two different disorders.[10]
A study of the volume of the gray and white matter in the anterior cingulate gyrus in patients with schizophrenia and their healthy first and second degree relatives revealed no significant difference in the volume of the white matter in the schizophrenic patients and their healthy relatives. Nonetheless a significant difference in the volume of gray matter was detected, schizophrenic patients had smaller volume of gray matter than their second degree relatives, but not relative to their first degree relatives. Both schizophrenic and their first degree healthy relatives have smaller gray matter volume than the second degree healthy relatives. It appears that genes are responsible for the decreased volume of gray matter in schizophrenic patients.[11]
Fujiwara et al. (2007) did an experiment in which they correlated the size of anterior cingulate gyrus in schizophrenic patients with their functioning on social cognition, psychopathology and emotions with control group. The smaller the size of anterior cingulate gyrus, the lower was the level of social functioning and the higher was the psychopathology in schizophrenic patient. The anterior cingulate gyrus was found to be bilaterally smaller in patients with schizophrenia as compared with control group. No difference in IQ tests and basic visuoperceptual ability with facial stimuli was found between schizophrenia patients and the control.[12]
Summary[]
Schizophrenic patients have differences in the anterior cingulate gyrus when compared with controls. Anterior cingulate gyrus was found to be smaller in schizophrenic patients.[12] The volume of the gray matter in the anterior cingulate gyrus was found to be lower in schizophrenic patients.[10][11] Healthy females have larger rostral anterior cingulate gyrus than males, this sex difference in size is absent in schizophrenic patients.[9] Metabolic rate of glucose was lower in the left anterior cingulate gyrus and in the right posterior cingulate gyrus.[10]
In addition to changes in the cingulate cortex more brain structures show changes in schizophrenic patients as compared to controls. The hippocampus in schizophrenic patients was found to be smaller in size when compared with controls of the same age group,[13] and, similarly, the caudate and putamen were found to be smaller in volume in a longitudinal study of schizophrenic patients.[14] While the volume of gray matter is smaller, the size of the lateral and third ventricles is larger in schizophrenic patients.[15]
| Anterior cingulate cortex
]]Functioning[]
Difficult dilemmas tend to elicit increased activity in the anterior cingulate cortex, a brain region associated with "response conflict." And when people make utilitarian judgments in response to these difficult dilemmas, Joshua Greene has found they exhibit increased activity in anterior regions of the dorsolateral prefrontal cortex.
See also[]
References[]
- ↑ DOI:10.1523/JNEUROSCI.4874-09.2010
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ McFarland, N. R., Harber, S. N. (2000). Convergent Inputs from Thalamic Motor Nuclei and Frontal Cortical Areas to the Dorsal Striatum in the Primate. The Journal of Neuroscience, May 15, 2000, 20(10):3798–3813.
- ↑ 3.0 3.1 DOI:10.1002/hipo.450020102
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ 4.0 4.1 4.2 DOI:10.1007/BF00237497
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ DOI:10.1016/0304-3940(83)90066-6
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ Petrides, M, Pandya, DN (2007 Oct 24). Efferent association pathways from the rostral prefrontal cortex in the macaque monkey.. The Journal of neuroscience : the official journal of the Society for Neuroscience 27 (43): 11573-86.
- ↑ 7.0 7.1 DOI:10.1002/cne.902870402
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ DOI:10.1093/brain/74.2.233
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ 9.0 9.1 DOI:10.1016/S0006-3223(02)01483-X
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ 10.0 10.1 10.2 DOI:10.1016/j.schres.2004.02.025
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ 11.0 11.1 DOI:10.1016/j.schres.2010.06.014
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ 12.0 12.1 DOI:10.1016/j.neuroimage.2007.03.068
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ DOI:10.4088/JCP.08m04574yel
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ DOI:10.1016/j.schres.2009.06.022
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand - ↑ DOI:10.1016/j.schres.2010.03.036
This citation will be automatically completed in the next few minutes. You can jump the queue or expand by hand
Further reading[]
- Vogt BA, Rosene DL, Pandya DN, Science 204:205+
External links[]
- Four Regions of Cingulate Cortex and Disease Vulnerability, Brent A. Vogt.
- BrainMaps at UCDavis Cingulate
- Mapping 'self' and 'other' in the brain
| This page uses Creative Commons Licensed content from Wikipedia (view authors). |